
Você já perdeu flores ou vagens da soja sem encontrar sintomas visuais que justificassem a perda? Solo corrigido, nutrição ajustada, sem pragas aparentes. Ainda assim, a planta descartou estruturas reprodutivas que exigiram energia para se formar — e o campo não deu nenhum aviso antes.
Esse tipo de situação é mais comum do que parece e, na maioria das vezes, está ligado a decisões fisiológicas internas da planta, mediadas por sinalização hormonal.
O etileno é o único fitormônio gasoso entre os principais reguladores vegetais. Por se difundir livremente pelos espaços intercelulares, sem depender de transportadores de membrana, ele atravessa tecidos em poucos minutos.
Essa velocidade permite que um tecido sob estresse sinalize para tecidos vizinhos antes mesmo de o problema se tornar visível. Em frutos armazenados, um único fruto danificado pode acelerar a senescência dos vizinhos em 30 a 50%, porque o etileno liberado se difunde rapidamente pelo ambiente (Zhang et al., 2020).
No campo, embora parte do hormônio se dissipe na atmosfera, sua atuação continua sendo decisiva. O etileno:
- regula a abscisão floral em soja entre R1 e R5;
- coordena a formação de aerênquima em milho sob alagamento;
- sincroniza a desfolha em algodão antes da colheita mecânica;
- participa da indução da antese em café após ciclos de déficit hídrico.
Ou seja, ele atua diretamente na definição de produtividade — muitas vezes antes que qualquer sinal visível apareça.
Este artigo apresenta a biossíntese do etileno, a lógica de percepção e sinalização, as respostas em soja, milho, algodão e café, os novos conhecimentos envolvendo nitrogênio, nodulação e dano mecânico, e as ferramentas disponíveis para interferir nessa rota fisiológica.
O que torna o etileno diferente dos outros fitormônios
Auxinas, giberelinas e citocininas precisam de transportadores ativos para se mover célula a célula. O etileno não. Por ser gasoso à temperatura ambiente, ele se difunde pelos espaços intercelulares e atravessa membranas com facilidade, sem depender de nenhum sistema de transporte (Figura 1).
Essa característica possui implicação agronômica prática direta. Um tecido sob estresse pode sinalizar rapidamente para tecidos adjacentes em minutos, muito antes de o problema se manifestar visualmente.
Em frutos armazenados com ventilação inadequada, o etileno produzido por um fruto danificado se acumula no ambiente e acelera o amadurecimento dos demais.
Em solos alagados, onde a difusão de gases é restrita, o etileno se acumula nos tecidos e ativa respostas que a planta não ativaria em solo bem drenado.
Em campo aberto, parte significativa do hormônio produzido se dissipa rapidamente. Isso significa que a intensidade da resposta, duração e alcance da resposta dependem diretamente do ambiente ao redor da planta.
De maneira prática, o etileno raramente age de forma localizada. Sua produção em um tecido pode modificar o comportamento fisiológico de regiões distantes da planta. Reconhecer esse caráter sistêmico é fundamental para interpretar sintomas que aparecem longe da causa original.

Como a planta produz etileno: a rota biossintética e o ciclo de Yang
A biossíntese do etileno segue três conversões sequenciais:
- A metionina é convertida em S-adenosilmetionina (SAM) pela SAM sintetase.
- Em seguida, o SAM é convertido em ácido 1-aminociclopropano-1-carboxílico (ACC) pela ACC sintase (ACS).
- Por fim, o ACC é oxidado a etileno pela ACC oxidase (ACO), reação que libera também CO₂ e cianeto de hidrogênio (HCN).
Rota resumida:
Metionina → SAM → ACC → etileno

O principal ponto de controle da rota está na etapa catalisada pela ACC sintase, responsável por definir a taxa efetiva de produção do hormônio (Pattyn et al., 2021).
Estresses abióticos como seca, alagamento, salinidade e temperatura extrema aumentam a expressão de genes que codificam ACS.
Em algodão, Li et al. (2022) identificaram múltiplos genes ACS responsivos a estresses abióticos, com padrões de expressão específicos para cada tipo de estresse.
A ACO depende de oxigênio para converter ACC em etileno. Em solos alagados, onde o O₂ difunde lentamente, a ACO tem atividade reduzida nas raízes. O ACC produzido se acumula localmente e é transportado para as partes aéreas.
Nas folhas e caules, com disponibilidade de O₂, a ACO converte o precursor acumulado em etileno. Esse mecanismo explica por que plantas alagadas produzem etileno nas partes aéreas mesmo com raízes em hipóxia.
A planta não esgota o pool de metionina durante a produção de etileno porque recicla o subproduto gerado. Quando o SAM é convertido em ACC, a reação libera 5’-metiltioadenosina (MTA). O ciclo de Yang converte a MTA de volta à metionina via sequência de reações enzimáticas. Esse ciclo sustenta a produção contínua de etileno em situações de alta demanda.
López et al. (2022) mostraram que o ACC pode atuar como molécula sinalizadora em cafeeiros com efeitos sobre a antese independentes da conversão final em etileno. O ACC não é apenas precursor. Essa descoberta reposiciona o entendimento da rota: a biossíntese tem múltiplos pontos de controle.
Como a planta percebe o etileno: receptores, cascata e diferença entre cultivares
A percepção do etileno opera por regulação negativa.
Os receptores localizados na membrana do retículo endoplasmático funcionam como repressores ativos na ausência do hormônio. Quando não há etileno, eles mantêm a cascata de sinalização bloqueada. Quando o etileno se liga aos receptores, inativa os repressores e libera a resposta gênica.
Binder (2020) descreve cinco receptores de etileno em Arabidopsis divididos em duas subfamílias.
- Subfamília I: ETR1 e ERS1.
- Subfamília II: ETR2, ERS2 e EIN4.
Todos requerem um átomo de cobre para ligar o etileno. Sem disponibilidade de cobre nos tecidos, a percepção do hormônio fica comprometida independentemente da quantidade produzida.
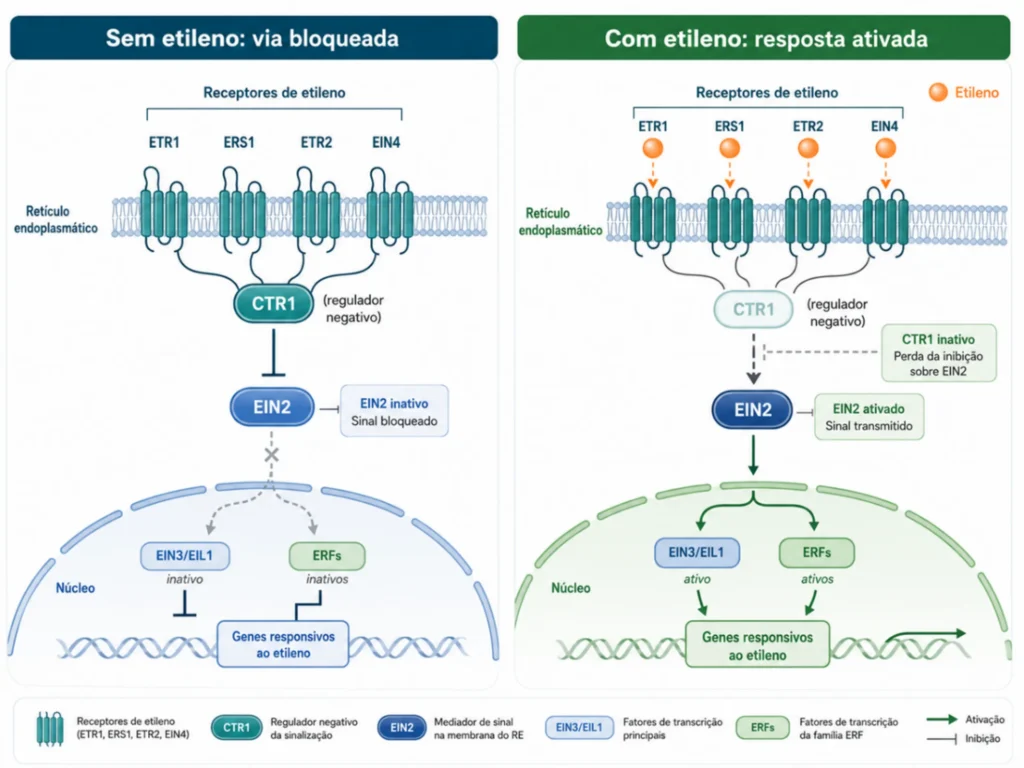
A cascata envolve as proteínas CTR1, EIN2, EIN3 e fatores de transcrição da família ERF (Ethylene Response Factor). Na ausência de etileno, CTR1 fosforila EIN2 mantendo-o inativo.
A ligação do etileno inativa os receptores, CTR1 para de fosforilar EIN2, e o domínio EIN2-CEND é clivado e transloca-se ao núcleo. Park et al. (2023) mostraram que o próprio CTR1 também migra do retículo endoplasmático ao núcleo em resposta ao etileno, refinando o modelo clássico.
A implicação agronômica direta é esta: duas cultivares podem produzir o mesmo nível de etileno e responder de formas completamente diferentes.
A sensibilidade dos receptores e a eficiência da cascata variam entre genótipos. Wang et al. (2022) identificaram em milho o fator de transcrição ZmERF21, cuja superexpressão aumentou a tolerância à seca ao modular a sensibilidade ao etileno.
Etileno e estresses abióticos
Fatma et al. (2022) revisaram o papel do etileno em múltiplos estresses e identificaram um padrão consistente: a resposta pode ser adaptativa ou destrutiva dependendo da cultura, do estádio fenológico, da intensidade do estresse e do tecido. Não existe uma resposta única e universal.
Alagamento e hipóxia em milho e soja
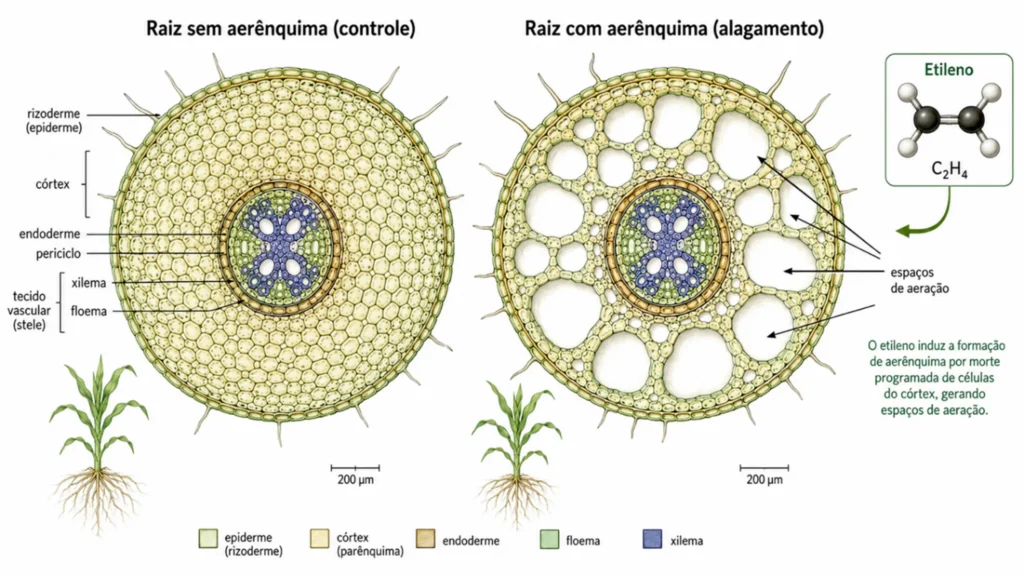
A Embrapa Milho e Sorgo desenvolveu a cultivar Saracura BRS 4154 após 18 ciclos de seleção para tolerância ao alagamento.
Concenço et al. (2022) mostraram que essa cultivar mantém maior atividade de álcool desidrogenase e menor acúmulo de acetaldeído sob hipóxia.
O etileno acumulado nas raízes inundadas desencadeia a formação de aerênquima, tecido com grandes espaços intercelulares que facilita a difusão de O₂ da parte aérea para as raízes.
Kim et al. (2015) compararam linhas de soja com diferente tolerância ao alagamento e encontraram padrões hormonais distintos entre tolerantes e suscetíveis.
Posteriormente, Kim et al. (2018) testaram a aplicação exógena de etileno em soja alagada e registraram indução de raízes adventícias, hipertrofia de lenticelas e aumento da expressão de glutationa-S-transferases. O etileno, nesse contexto, coordena a reorganização fisiológica para sobrevivência.
Déficit hídrico em milho e soja
Shi et al. (2017) usaram CRISPR-Cas9 para inserir o promotor GOS2 a montante do gene ARGOS8 em milho. O ARGOS8 codifica um regulador negativo da resposta ao etileno. Com sua superexpressão, a planta responde menos intensamente ao hormônio sob estresse, com ganho de produtividade em condições de estresse no florescimento.
Em soja, a tecnologia HB4 usa um gene de girassol que reduz a sensibilidade ao etileno sob déficit hídrico moderado.
Arraes et al. (2015) mostraram que cultivares tolerantes à seca não eliminam a produção de etileno. Eles modulam a intensidade e o tempo da resposta para que ela seja adaptativa, não destrutiva.
Salinidade e crosstalk com poliaminas e óxido nítrico
Gill e Tuteja (2010) revisaram a interação entre etileno, poliaminas e óxido nítrico em plantas sob estresse salino. Poliaminas como putrescina, espermidina e espermina estabilizam membranas, reduzem a produção de espécies reativas de oxigênio e modulam genes de resposta a estresse.
Khan et al. (2024) atualizaram essa análise, mostrando que a interação entre etileno, poliaminas e NO é dependente de contexto.
O SAM é disputado entre a rota do etileno e a rota das poliaminas. Quando a célula investe SAM em poliaminas, reduz a disponibilidade para etileno. Esse balanço define se a resposta ao estresse será protetora ou acelerada em direção à senescência.
Etileno e eficiência do uso de nitrogênio
O etileno regula a arquitetura das raízes laterais sob baixa disponibilidade de nitrogênio, impactando diretamente a exploração do solo e a capacidade de captura de nutrientes. Em solos tropicais com alta variabilidade espacial de N, essa regulação tem implicação direta na eficiência do uso de nitrogênio (NUE).
Quando o N disponível cai abaixo de um limiar, o etileno inibe o alongamento de raízes laterais, reduzindo a exploração do perfil do solo.
Cultivares com menor sensibilidade ao etileno nas raízes mantêm melhor arquitetura radicular sob baixo N, capturam maior volume de nitrogênio disponível e respondem melhor ao N aplicado via adubação.
A NUE, portanto, não depende apenas da dose aplicada. Depende também da capacidade da planta de ajustar o sistema radicular via sinalização por etileno.
Etileno e nodulação em soja
Guinel e Geil (2002) revisaram a relação entre etileno e nodulação em leguminosas e mostraram que o etileno endógeno suprime a infecção por Bradyrhizobium japonicum. Mutantes insensíveis ao etileno formam mais nódulos do que plantas selvagens. Picos de etileno gerados por estresse precoce durante V1-V4 podem reduzir drasticamente o número de nódulos formados.
Compactação, alagamento temporário e déficit hídrico nas primeiras semanas após emergência aumentam a produção de etileno nas raízes e comprometem a nodulação da safra. A fixação biológica de nitrogênio pode suprir 50 a 70% da demanda de N da soja. Perder essa contribuição por estresse precoce é uma questão econômica direta.
Atenção técnica: quando a nodulação está fraca sem falha aparente do inoculante, investigue estresses precoces. O problema pode estar no etileno gerado antes mesmo de o inoculante ter chance de agir.
Etileno, abscisão e tomada de decisão
A abscisão é uma decisão coordenada, não um acidente. A planta avalia o status energético, o suprimento de fotoassimilados e o balanço hormonal antes de ativar a zona de abscisão.
O etileno coordena esse processo ao induzir as enzimas que degradam componentes da parede celular na região de separação.
O sintoma que você observa no campo (a flor caída ou a vagem no chão) é a etapa final de um processo iniciado dias antes. A intervenção mais eficaz acontece antes da janela crítica, não depois que a perda já ocorreu.
Abortamento floral em soja
A soja produz mais flores do que consegue sustentar. Apenas 20 a 40% das flores resultam em vagens maduras. A janela crítica é R1 a R5.
Cheng et al. (2013) analisaram o transcriptoma de órgãos reprodutivos em soja e identificaram genes mediados por etileno associados ao desenvolvimento e à abscisão.
Quando estresses durante R1 a R5 reduzem a disponibilidade de fotoassimilados, o etileno coordena o descarte das estruturas que não estão recebendo suprimento adequado.
Déficit hídrico, temperatura alta, sombreamento e deficiência nutricional ativam essa cascata. Quando o abortamento é visível, a decisão fisiológica já foi tomada.

Abscisão em algodão
Chen et al. (2022) identificaram o gene GhArfGAP em algodão, com expressão na camada de abscisão de maçãs tratadas com etileno. Esse gene regula o tráfego vesicular no Golgi e a reorganização da parede celular durante a separação.
Em algodão, apenas 24 a 36% das flores resultam em capulhos maduros (Chen et al., 2014).
O ethephon é usado como desfolhante antes da colheita mecânica, acelerando a abscisão foliar e reduzindo a contaminação da fibra. Liao et al. (2023) compararam thidiazuron e ethephon como desfolhantes e mostraram que ambos aumentam a produção de etileno, com padrões de expressão gênica distintos.
A escolha entre eles depende do estádio, das condições ambientais e do objetivo da aplicação.
Regeneração de tecidos e formação da periderme após dano mecânico
Quando raízes, tubérculos ou qualquer órgão vegetal sofrem dano mecânico, a planta precisa selar a lesão antes que patógenos entrem. O processo chama-se suberização e envolve a formação de uma camada de células denominada periderme. O etileno e o oxigênio controlam em conjunto o início e o encerramento desse processo.
O dano mecânico induz uma onda local de etileno em menos de duas horas. Esse sinal recruta células cambiais ao redor da lesão para diferenciação em periderme.
A disponibilidade de O₂ regula a duração da resposta: sem aeração adequada, como em tubérculos armazenados com ventilação insuficiente, a suberização fica incompleta mesmo com etileno elevado.
Para raízes, tubérculos e cana após corte mecânico, controlar a ventilação no armazenamento não é apenas questão de temperatura.
O mesmo princípio se aplica a granizo, poda em frutíferas e ataque de insetos mastigadores em campo. Cada dano gera um pico local de etileno, e a qualidade da cicatrização depende do O₂ disponível no tecido adjacente.
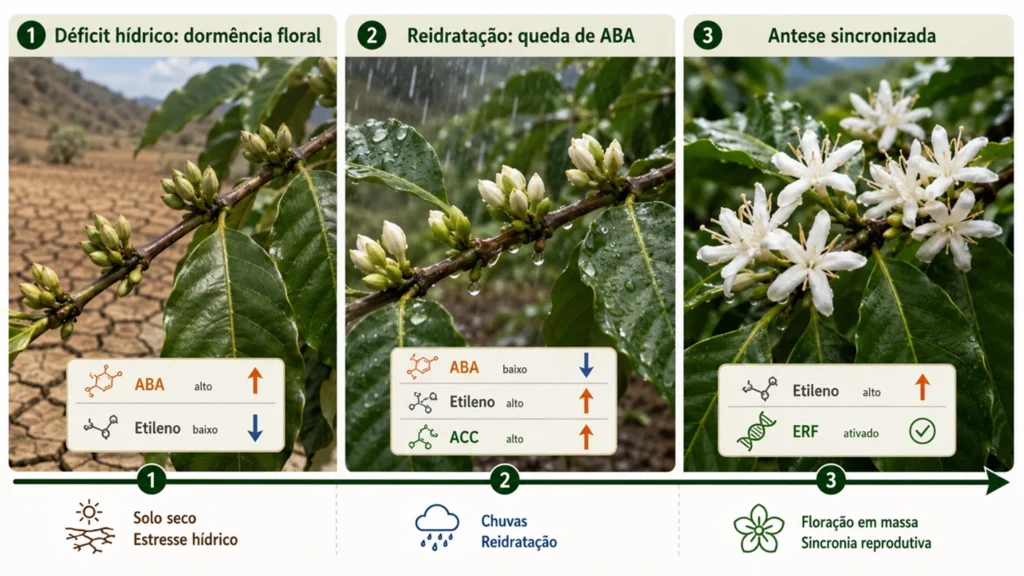
Maturação e antese em café
López et al. (2021) estudaram a maturação de frutos de café sob déficit hídrico e mostraram que o estresse aumenta a produção de etileno e acelera a maturação. Ságio et al. (2014) demonstraram que o etileno regula a antese de flores de café, coordenando a abertura floral após períodos de restrição hídrica.
López et al. (2022) mostraram que o ACC acumula em botões florais e atua como sinal independente do etileno, modulando a resposta ao ABA. Confirmaram, ainda que a interação entre etileno e ABA regula a antese, com implicações diretas para a sincronia de floração e a uniformidade de maturação.
Interação do etileno com outros hormônios
O etileno raramente age em isolamento. A resposta final depende do contexto hormonal do tecido. As quatro interações abaixo são as mais documentadas com aplicação agronômica direta.
Etileno e auxina
Beyer (1973) demonstrou que o etileno modifica o transporte de auxina, reduzindo a concentração de auxina na zona de abscisão.
Suttle et al. (1991) mostraram em algodão que o etileno reduz os níveis de auxina livre (IAA) em cerca de 55% e inibe o transporte basípeto de IAA em pecíolos. A sensibilidade à auxina varia com a idade do tecido, e a interação entre os dois hormônios define o momento e a intensidade da abscisão.
Etileno e ABA
Antonietta et al. (2023) mostraram em soja que a exposição prolongada a baixa temperatura dessensibiliza o fechamento estomático induzido por ABA via processo dependente de etileno. Em café, López et al. (2022) demonstraram que durante o déficit hídrico o ABA mantém a dormência floral. Com a reidratação, a queda de ABA e o aumento de etileno sincronizam a abertura floral.
Etileno e giberelina
Em plantas inundadas, o etileno acumulado nos tecidos induz a síntese de giberelina. A giberelina promove o alongamento de entrenós, mecanismo de fuga que eleva as folhas acima da superfície da água.
Em milho e soja, essa resposta é menos pronunciada que em espécies de áreas alagáveis. A tolerância ao alagamento nessas culturas depende mais da formação de aerênquima.
Etileno e citocinina
Citocininas retardam a senescência foliar. O etileno a acelera. O balanço entre os dois hormônios determina a duração da área foliar verde e a eficiência de enchimento de grãos. Em milho, cultivares do tipo stay-green têm maior relação citocinina/etileno durante o enchimento e mantêm fotossíntese ativa por mais tempo, resultando em maior peso de grãos.
A tabela abaixo resume as principais interações hormonais e suas implicações agronômicas:
Tabela 1. Principais interações hormonais e suas implicações agronômicas
| Hormônio | Tipo de interação | Implicação agronômica |
| Auxina | Antagonismo no transporte e percepção | Controle da abscisão em algodão e soja |
| ABA | Sinergismo em estresse hídrico; antagonismo na antese | Resposta à seca em soja; sincronização floral em café |
| Giberelina | Sinergismo em fuga de alagamento | Alongamento de entrenós em plantas inundadas |
| Citocinina | Antagonismo na senescência | Stay-green em milho; enchimento de grãos |
| Poliaminas | Competição por SAM como precursor | Resposta ao estresse salino e oxidativo |
| Óxido nítrico | Crosstalk dependente de contexto | Modulação da resposta a estresses combinados |
Ethephon, 1-MCP e inibidores: ferramentas de manejo da rota do etileno
O conhecimento sobre biossíntese e percepção do etileno gerou ferramentas que interferem em pontos específicos da rota.
Cada ferramenta tem mecanismo, aplicação e limitação bem definidos. A escolha depende do objetivo: estimular ou bloquear a resposta.
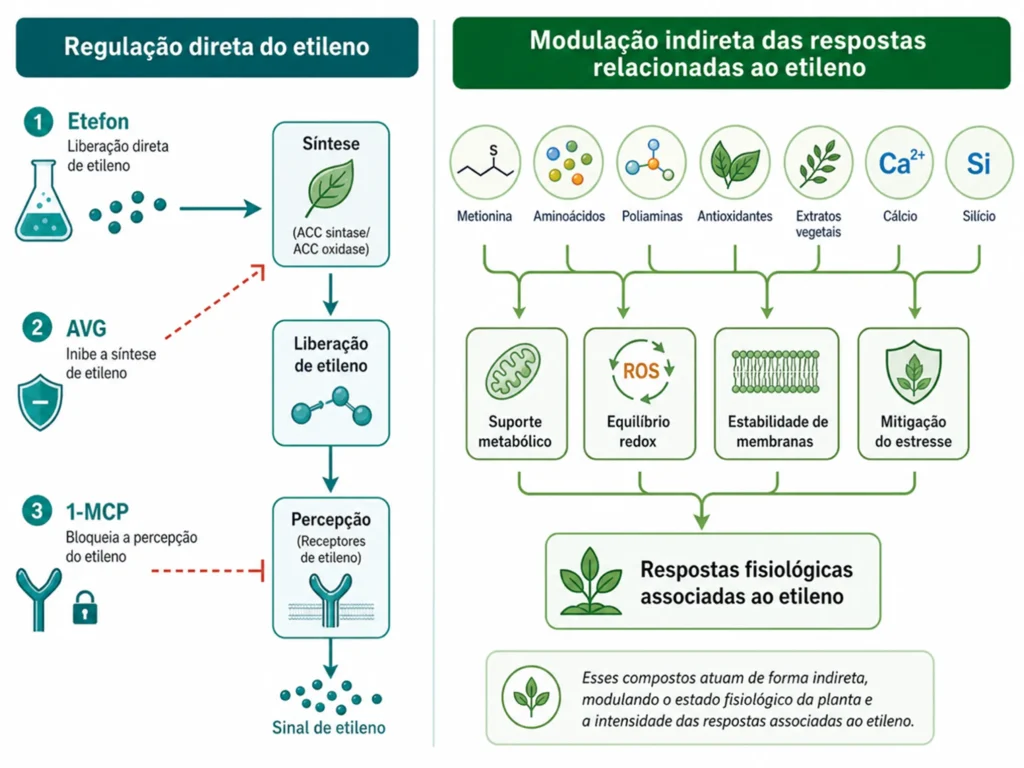
Ethephon: liberação de etileno nos tecidos
O ethephon (ácido 2-cloroetilfosfônico) libera etileno após penetrar nos tecidos. O pH alcalino do citosol promove sua decomposição, liberando etileno, cloreto e fosfato.
Em algodão, é aplicado como desfolhante antes da colheita mecânica, com dose típica de 0,5 a 1,5 L ha⁻¹. Aplicações acima de 30°C aumentam a taxa de liberação e podem causar desfolha excessiva.
Em café, uniformiza a maturação de frutos em pré-colheita. Em cana, aumenta o teor de sacarose nos colmos. Em abacaxi, sincroniza a floração para colheita programada. O produto deve ser usado conforme registro, estádio fenológico e condições ambientais.
AVG e cobalto: bloqueio da síntese
O AVG (aminoetoxivinilglicina) inibe a ACC sintase, bloqueando a conversão de SAM em ACC. Seu uso é mais comum em fruticultura para retardar a maturação e prolongar a vida pós-colheita. O cobalto (Co²⁺) também inibe a ACC sintase por competição com o cofator piridoxal fosfato. O uso de cobalto é restrito em vários países por razões de segurança ambiental.
1-MCP: bloqueio dos receptores
O 1-metilciclopropeno (1-MCP) se liga aos receptores de etileno com afinidade dez vezes maior que o próprio hormônio, ocupando o sítio de ligação sem ativar a cascata.
Zhang et al. (2020) conduziram meta-análise com 44 indicadores de amadurecimento e registraram redução média de 65% na produção de etileno e redução de 20 dos 44 indicadores em pelo menos 22%.
Em algodão, Chen et al. (2014) mostraram que o 1-MCP aumentou o peso de capulhos aos 20 dias após a floração e aumentou a eficiência fotossintética em folhas subtendentes.
Djanaguiraman et al. (2010) mostraram em soja que o 1-MCP reduziu a senescência foliar prematura causada por estresse térmico, resultando em maior fixação de sementes. LÓPEZ et al. (2022) mostraram em café que o 1-MCP pode induzir antese ao bloquear receptores fora da janela adequada.
Dano mecânico e pico de etileno na logística pós-colheita
Cada impacto físico no fruto é um evento fisiológico. O dano mecânico ativa a ACC sintase via MAP quinases em menos de duas horas. O ACC produzido é convertido em etileno pela ACO e difunde para os frutos adjacentes. Um fruto danificado pode acelerar a senescência dos vizinhos em 30 a 50% (Zhang et al., 2020).
O 1-MCP bloqueia o receptor, mas não elimina o etileno produzido. O controle da causa é anterior ao produto. Ele está na regulagem de colhedoras, na altura de queda em esteiras, no tipo de embalagem e na velocidade de transporte.
Reduzir dano mecânico é reduzir produção endógena de etileno. Essa é a intervenção mais eficiente antes de qualquer produto.
Tiossulfato de prata e nitrato de prata
Os íons prata (Ag⁺) se ligam ao sítio de cobre dos receptores de etileno e os inativam.
O tiossulfato de prata (STS) e o nitrato de prata são usados principalmente em cultura de tecidos e pós-colheita de flores de corte.
O uso em campo é restrito por razões regulatórias e de segurança ambiental.
Compostos naturais e modulação indireta do etileno
Compostos naturais não bloqueiam o etileno de forma direta.
O argumento técnico correto é que eles melhoram o estado fisiológico da planta e reduzem a necessidade de respostas de emergência associadas ao etileno.
Essa distinção protege o produtor de expectativas sem base e direciona o uso para onde há evidência.
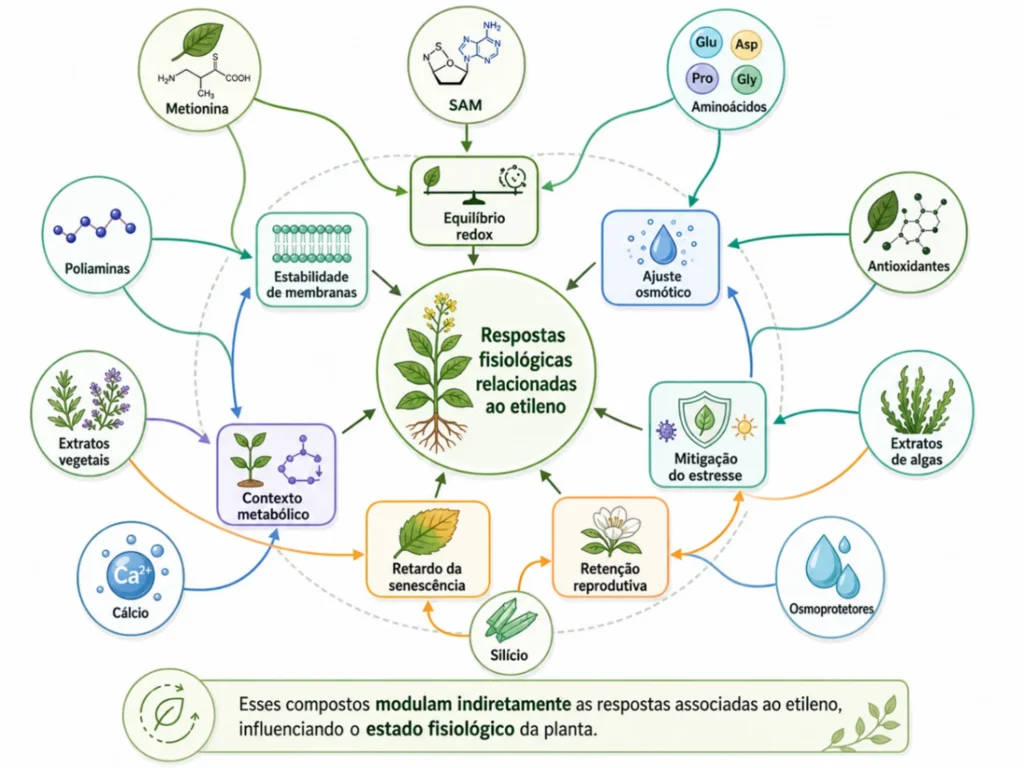
Metionina: precursor, não regulador direto
A metionina é o ponto de partida da biossíntese do etileno. Isso não significa que aplicá-la exogenamente aumente a produção do hormônio. O passo limitante é a ACC sintase, não a disponibilidade do precursor. A planta recicla a metionina pelo ciclo de Yang.
O fornecimento de aminoácidos contendo enxofre pode dar suporte ao metabolismo vegetal em situações de deficiência. O efeito sobre a produção de etileno é dependente de contexto e não deve ser generalizado sem evidência experimental específica.
Poliaminas, antioxidantes e equilíbrio redox
Hasan et al. (2021) revisaram o papel de poliaminas em células sob estresse e mostraram que espermina e espermidina estabilizam membranas, reduzem a produção de espécies reativas de oxigênio e modulam genes de resposta a estresse.
Quando a célula investe SAM em poliaminas, há menos precursor disponível para a rota do etileno.
Compostos fenólicos, flavonoides e nutrientes cofatores de enzimas antioxidantes seguem a mesma lógica. Menos dano oxidativo significa menos sinalização de emergência e menor indução de etileno por degradação celular. O efeito é indireto, mas documentado por Gill e Tuteja (2010).
Cálcio, boro e integridade estrutural
Cálcio e boro têm função estrutural na parede celular e nas membranas. O cálcio estabiliza pectinas.
O boro forma complexos com polissacarídeos, aumentando a rigidez da parede. Tecidos com maior integridade estrutural são menos vulneráveis ao colapso celular que desencadeia respostas de emergência.
Em estruturas reprodutivas, paredes celulares mais estáveis reduzem a sensibilidade à formação de zonas de abscisão. Manejar cálcio e boro em fases críticas não é bloquear o etileno. É reduzir os sinais que disparam sua produção excessiva.
Como interpretar o etileno no campo
A tabela 2 organiza situações observáveis no campo, a relação fisiológica com o etileno e o próximo passo técnico. Não é diagnóstico fechado. O campo é multifatorial e o etileno quase sempre age junto com outros sinais hormonais e ambientais.
Tabela 2. Situações observada no campo, a relação fisiológica com o etileno e o próximo passo técnico
| Situação observada | Relação com etileno | Próximo passo técnico |
| Solo encharcado por mais de 48 h | Hipóxia radicular aumenta produção; acúmulo por difusão restrita | Avaliar drenagem; monitorar arquitetura radicular |
| Abortamento floral em soja R1-R5 | Estresse ativa zonas de abscisão via etileno | Identificar estressor antes de R1 |
| Nodulação fraca sem falha do inoculante | Pico de etileno por estresse precoce suprime Bradyrhizobium | Investigar V1-V4: compactação, alagamento, déficit |
| Senescência foliar antecipada | Balanço etileno/CK desfavorável; remobilização acelerada | Avaliar N, Mg e estado hídrico foliar |
| Maturação irregular em café | Diferença de sensibilidade ao etileno entre frutos e ramos | Considerar uniformizadores de maturação registrados |
| Desfolha excessiva após ethephon | Dose alta ou temperatura acima de 30°C acelerou liberação | Ajustar dose; evitar aplicação em altas temperaturas |
| Pós-colheita com amadurecimento acelerado | Dano mecânico ativou ACC sintase; difusão para vizinhos | Reduzir dano na colheita e logística; avaliar 1-MCP |
| Deficiência de N com raízes laterais curtas | Etileno inibe alongamento radicular sob baixo N | Avaliar N no perfil; ajustar adubação de base |
Conclusão
O etileno coordena decisões fisiológicas sob pressão em soja, milho, algodão e café.
Na soja, regula quantas flores vão se tornar vagens e se a nodulação vai se estabelecer ou ser suprimida por estresse precoce.
No milho alagado, define se a planta forma aerênquima e sobrevive ou entra em colapso metabólico.
No algodão, é a base dos protocolos de desfolha e maturação para colheita mecânica. No café, coordena a antese sincronizada e o amadurecimento uniforme dos frutos.
A biossíntese via metionina, SAM, ACC e etileno, o ciclo de Yang que sustenta essa produção, a cascata de receptores que transforma o gás em resposta gênica e as ferramentas que interferem nesses pontos formam um conjunto de conhecimentos com aplicação direta no diagnóstico de campo.
A pergunta que você aprende a fazer muda a forma de ler a lavoura: onde a planta está nessa rota, o que a levou até ali e qual a consequência agronômica?
Você observa abortamento floral em R3.
O que disparou a produção de etileno? Déficit hídrico? Temperatura alta? Nodulação comprometida por estresse precoce?
Você identifica a causa, intervém antes da perda aparecer e protege a produtividade com base na fisiologia.
O etileno nas plantas não é um problema a eliminar. É um sinal a interpretar.
—
Quer saber mais sobre hormônios? Baixe o infográfico sobre hormônios vegetais: Importância e interações na planta. Clique em Baixe agora!

E se você quer saber mais sobre fisiologia e nutrição de plantas, conheça a Pós-Graduação em Fisiologia e Nutrição de Plantas da AgroAdvance e aprofunde seu conhecimento sobre os mecanismos fisiológicos das plantas para gerar resultados no campo.

Referências
ANTONIETTA, M. et al. Prolonged low temperature exposure de-sensitises ABA-induced stomatal closure in soybean, involving an ethylene-dependent process. Plant Cell and Environment, v. 46, n. 8, p. 2537-2551, 2023. DOI: 10.1111/pce.14590.
ARRAES, F. B. M. et al. Implications of ethylene biosynthesis and signaling in soybean drought stress tolerance. BMC Plant Biology, v. 15, n. 1, p. 213, 2015. DOI: 10.1186/S12870-015-0597-Z.
BEYER, E. M. Abscission: support for a role of ethylene modification of auxin transport. Plant Physiology, v. 52, n. 1, p. 1-5, 1973. DOI: 10.1104/PP.52.1.1.
BINDER, B. M. Ethylene signaling in plants. Journal of Biological Chemistry, v. 295, n. 22, p. 7710-7725, 2020. DOI: 10.1074/jbc.REV120.010854.
CHEN, D. et al. Effect of 1-MCP on boll development and subtending leaves of cotton (Gossypium hirsutum L.) plants. American Journal of Plant Sciences, v. 5, n. 21, p. 3186-3197, 2014. DOI: 10.4236/AJPS.2014.521350.
CHEN, Y. et al. Cloning and Bioinformatics Analysis of GhArfGAP in Cotton (Gossypium hirsutum) Boll Abscission Layer With Ethylene Treatment. Frontiers in Plant Science, v. 13, p. 841161, 2022. DOI: 10.3389/fpls.2022.841161.
CHENG, W. H. et al. RNA-seq analysis reveals ethylene-mediated reproductive organ development and abscission in soybean (Glycine max L. Merr.). Plant Molecular Biology Reporter, v. 31, n. 3, p. 607-619, 2013. DOI: 10.1007/S11105-012-0533-4.
DJANAGUIRAMAN, M. et al. Ethylene production under high temperature stress causes premature leaf senescence in soybean. Functional Plant Biology, v. 37, n. 11, p. 1071-1084, 2010. DOI: 10.1071/FP10089.
FATMA, M. et al. Ethylene and sulfur coordinately modulate the antioxidant system and ABA accumulation in mustard plants under salt stress. Plants, v. 11, n. 17, p. 2211, 2022. DOI: 10.3390/plants11172211.
GILL, S. S.; TUTEJA, N. Polyamines and abiotic stress tolerance in plants. Plant Signaling & Behavior, v. 5, n. 1, p. 26-33, 2010. DOI: 10.4161/psb.5.1.10291.
GUINEL, F. C.; GEIL, R. D. A model for the development of the rhizobial and arbuscular mycorrhizal symbioses in legumes and its use to understand the roles of ethylene in the establishment of these two symbioses. Canadian Journal of Botany, v. 80, n. 7, p. 695-720, 2002.
ALCÁZAR, R. et al. Polyamines: small molecules with large effects on plant abiotic stress tolerance. Cells, v. 10, n. 2, p. 261, 2021. DOI: 10.3390/cells10020261.
AHMAD, P. et al. Polyamines: role in plants under abiotic stress. In: IQBAL, N. et al. (Eds.). Plant Stress Mitigators. Academic Press, 2024. p. 227-247. 10.1007/978-94-007-4116-4_19,
KIM Y-H, HWANG S-J, WAQAS M, KHAN AL, LEE J-H, LEE J-D, NGUYEN HT AND LEE I-J (2015) Comparative analysis of endogenous hormones level in two soybean (Glycine max L.) lines differing in waterlogging tolerance. Front. Plant Sci. 6:714. doi: 10.3389/fpls.2015.00714
KIM Y, SEO CW, KHAN AL, MUN BG, SHAHZAD R, KO JW, YUN BW, PARK SK, LEE IJ. Exo-ethylene application mitigates waterlogging stress in soybean (Glycine max L.). BMC Plant Biol. 2018 Oct 22;18(1):254. doi: 10.1186/s12870-018-1457-4.
LI, Y. et al. Potential roles of 1-aminocyclopropane-1-carboxylic acid synthase genes in the response of Gossypium species to abiotic stress by genome-wide identification and expression analysis. Plants, v. 11, n. 11, p. 1524, 2022. DOI: 10.3390/plants11111524.
LIAO, B. et al. Comparative physiological and transcriptomic mechanisms of defoliation in cotton in response to thidiazuron versus ethephon. International Journal of Molecular Sciences, v. 24, n. 8, p. 7590, 2023. DOI: 10.3390/ijms24087590.
LÓPEZ, J. et al. Ethylene and 1-aminocyclopropane-1-carboxylic acid (ACC) act as signals during the coffee floral bud development and anthesis. Frontiers in Plant Science, v. 13, p. 824948, 2022. DOI: 10.3389/fpls.2022.824948.
PARK, H.L., SEO, D.H., LEE, H.Y. ET AL. Ethylene-triggered subcellular trafficking of CTR1 enhances the response to ethylene gas. Nat Commun 14, 365 (2023). https://doi.org/10.1038/s41467-023-35975-6
PATTYN, J. et al. The regulation of ethylene biosynthesis: a complex multilevel control circuitry. New Phytologist, v. 229, n. 2, p. 707-720, 2021. DOI: 10.1111/nph.16873.
CONCENÇO, G.; BERNARDI, N. D. C.; MELO, T. S.; SILVA, L. B. X.; RIBEIRO, I. B.; THEISEN, G.; PARFITT, J. M. B. Efeito do encharcamento do solo sobre milho e alternativas de manejo em terras baixas. Pelotas: Embrapa Clima Temperado, 2022. 8 p. (Embrapa Clima Temperado. Comunicado Técnico, 388). Disponível em: https://www.infoteca.cnptia.embrapa.br/infoteca/bitstream/doc/1146999/1/CPACT-Comunicado-Tecnico-388.pdf
SÁGIO, S. A. et al. Identification and expression analysis of ethylene biosynthesis and signaling genes provides insights into the early and late coffee cultivars ripening pathway. Planta, v. 239, n. 5, p. 1087-1097, 2014. DOI: 10.1007/s00425-014-2026-1.
SHI, J. et al. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnology Journal, v. 15, n. 2, p. 207-216, 2017. DOI: 10.1111/pbi.12603.
SUTTLE, J. C. et al. Ethylene-induced leaf abscission in cotton seedlings: the physiological bases for age-dependent differences in sensitivity. Plant Physiology, v. 95, n. 1, p. 29-33, 1991. DOI: 10.1104/PP.95.1.29.
WANG, Z. et al. ZmERF21 directly regulates hormone signaling and stress-responsive gene expression to influence drought tolerance in maize seedlings. Plant, Cell & Environment, v. 45, n. 3, p. 675-694, 2022. DOI: 10.1111/pce.14243.
ZHANG, J. et al. Meta-analysis of the effects of 1-methylcyclopropene (1-MCP) treatment on climacteric fruit ripening. Horticulture Research, v. 7, n. 1, p. 1-14, 2020. DOI: 10.1038/s41438-020-00405-x.
Sobre o autor:

Como citar este artigo:
BUENO, D.R. Etileno nas plantas: como o hormônio regula respostas ao estresse, abscisão e produtividade. Blog Agroadvance. Publicado: 29 Mai. 2026. Disponível em: https://agroadvance.com.br/blog-hormonios-vegetais-etileno/. Data de acesso: 20 jul. 2026




