
O milho é uma das culturas mais eficientes do ponto de vista fisiológico, sendo capaz de produzir elevadas quantidades de biomassa e grãos mesmo sob condições de altas temperaturas e elevada radiação solar. Mas o que realmente explica essa alta eficiência? Seria apenas o metabolismo fotossintético C4 ou existe uma integração mais profunda entre os processos metabólicos da planta?
Estima-se que o milho necessite entre 20 e 25 kg de nitrogênio para produzir uma tonelada de grãos, evidenciando sua alta exigência nutricional. No entanto, mesmo com aplicações elevadas de fertilizantes nitrogenados, grande parte desse nitrogênio não é convertida em produtividade, sendo perdida por volatilização, lixiviação ou desnitrificação.
Nesse cenário, surge uma pergunta central: por que, mesmo com alta disponibilidade de nitrogênio, nem sempre há resposta proporcional em rendimento?
A resposta está na fisiologia vegetal. No milho, o metabolismo do carbono e do nitrogênio operam de maneira altamente integrada. O metabolismo C4, além de aumentar a eficiência fotossintética, fornece energia, poder redutor e esqueletos de carbono essenciais para a assimilação do nitrogênio, tornando esse processo dependente da coordenação metabólica da planta.
Dessa forma, compreender a relação entre o metabolismo fotossintético C4, assimilação do nitrogênio, dinâmica do nutriente do solo e manejo agronômico é fundamental para aumentar a eficiência do uso do nitrogênio e maximizar a produtividade do milho em sistemas agrícolas modernos.
A Figura 1 apresenta de forma simplificada como o metabolismo C4 se conecta à assimilação do nitrogênio no milho, evidenciando a relação entre produção de energia, geração de esqueletos de carbono e formação de compostos nitrogenados responsáveis pela construção da produtividade da cultura.

Figura 1. Integração entre metabolismo C₄ e assimilação do nitrogênio no milho. Legenda: 1. Fixação inicial do carbono. O bicarbonato (HCO₃⁻), derivado do CO₂, reage com o fosfoenolpiruvato (PEP) pela ação da enzima PEPCase (fosfoenolpiruvato carboxilase), formando malato (C₄) nas células do mesófilo. 2. Transporte do malato. O malato (C₄) é transportado das células do mesófilo para as células da bainha vascular. 3. Descarboxilação do malato Na mitocôndria das células da bainha vascular, a enzima NADP-málica converte o malato em piruvato (C₃), liberando CO₂ para o ciclo de Calvin-Benson. 4. Retorno do piruvato. O piruvato (C₃) retorna das células da bainha vascular para as células do mesófilo. 5. Regeneração do fosfoenolpiruvato (PEP). No cloroplasto das células do mesófilo, a enzima PPDK (piruvato fosfato diquinase) converte o piruvato novamente em fosfoenolpiruvato (PEP), reiniciando o ciclo C₄.
Metabolismo C4 e assimilação de nitrogênio no milho
O milho é uma espécie tropical classificada como metabolismo fotossintético C4, pois os primeiros compostos orgânicos formados durante a assimilação do carbono possuem quatro carbonos. Esse é um dos principais diferenciais fisiológicos da cultura, conferindo elevada eficiência fotossintética sob condições de altas temperaturas, elevada radiação e déficit hídrico moderado. (Sage & Zhu, 2011; Hatch, 1987; Ermakova et al., 2020).
No primeiro passo desse processo (figura 2), o CO₂ difunde pelos estômatos e adentra as células do mesofilo foliar, onde é rapidamente convertido em bicarbonato pela ação da enzima anidrase carbônica (Hatch & Burnell, 1990; Studer et al., 2014). Em seguida, esse bicarbonato passa a ser substrato da PEP carboxilase, que catalisa sua reação com o fosfoenolpiruvato (PEP), formando oxalacetato, o primeiro composto orgânico estável de quatro carbonos do metabolismo fotossintético C4 (Hatch, 1987; Furbank, 2016).

A partir desse ponto, o oxalacetato pode seguir diferentes rotas metabólicas. Uma delas leva à formação de malato pela ação da malato desidrogenase, utilizando NADPH produzido na etapa fotoquímica como poder redutor. Esse malato é então transportado para as células da bainha do feixe vascular, onde sofre descarboxilação, liberando CO₂ em alta concentração para ser utilizado pela Rubisco no ciclo de Calvin (Edwards et al., 2004; Furbank, 2011).
Na segunda via, o oxalacetato forma aspartato por meio da ação da aspartato aminotransferase, enzima que catalisa a aminotransferência entre oxalacetato e glutamato. O aspartato é então transferido para as células da bainha do feixe vascular, onde perde seu grupamento amino, retornando a oxalacetato e posteriormente a malato, que novamente sofre descarboxilação para liberar CO₂ à Rubisco. Técnicas de marcação isotópica e análise de fluxos metabólicos em milho indicam que aproximadamente 30 a 40% do carbono pode circular entre mesofilo e bainha por essa via metabólica (Wingler et al., 1999; Arrivault et al., 2017).
Neste ponto, percebe-se claramente a integração entre o metabolismo do carbono e o metabolismo do nitrogênio na cultura do milho. Ao formar aspartato, parte do carbono assimilado passa a carregar nitrogênio orgânico oriundo da assimilação do nitrato via metabolismo do nitrogênio.
Além disso, quando o aspartato perde seu grupamento amino nas células da bainha do feixe, ocorre reciclagem de glutamato, molécula central no metabolismo nitrogenado, que pode ser redistribuída pelo floema para síntese de novos aminoácidos ou incorporação em proteínas estruturais e metabólicas (Foyer et al., 2011; Nunes-Nesi et al., 2010; Bloom, 2015).
Esse mecanismo é conhecido como concentrador de carbono e garante às espécies C4 elevada eficiência de carboxilação pela Rubisco.
Ao liberar CO₂ diretamente nas células da bainha do feixe vascular, há maior saturação dos sítios catalíticos da enzima, favorecendo sua atividade carboxilase e reduzindo sua atividade oxigenase, o que diminui a intensidade da fotorespiração e reduz perdas do carbono previamente assimilado (Sage, 2004; von Caemmerer & Furbank, 2016; Ghannoum et al., 2011; Ermakova et al., 2020).
Isso ajuda a explicar o rápido crescimento e a elevada capacidade produtiva do milho, especialmente sob condições de seca e altas temperaturas, onde espécies C4 apresentam vantagem fisiológica marcante sobre espécies C3 (Sage & Kubien, 2007; Yamori et al., 2014).
Em trabalho conduzido por Avila et al. (2016), avaliando quatro genótipos de milho sob irrigação normal e déficit hídrico severo, observou-se taxa fotossintética próxima de 35 μmol CO₂ m⁻² s⁻¹ em capacidade de campo e cerca de 15 μmol CO₂ m⁻² s⁻¹ sob seca intensa, redução de 57,14%. Em contraste, Campos et al. (2019), trabalhando com plantas de café (Coffea arabica), verificaram taxa fotossintética próxima de 12 μmol CO₂ m⁻² s⁻¹ em condições normais de irrigação e apenas 3 μmol CO₂ m⁻² s⁻¹ sob seca, redução de 75%.
Isso evidencia que, além de apresentar maior capacidade de assimilação de carbono em condições ideais, o milho também preserva melhor sua atividade fotossintética sob estresse hídrico, mantendo maior produção de açúcares e maior suporte metabólico ao crescimento.
Para sustentar essa elevada eficiência fotossintética, o milho apresenta alta exigência em nitrogênio, uma vez que grande parte de sua maquinaria metabólica é constituída por moléculas nitrogenadas (Evans, 1989; Sinclair & Horie, 1989; Mu et al., 2016).
Entre elas estão as clorofilas, proteínas do sistema fotoquímico de transporte de elétrons, enzimas do mecanismo concentrador de carbono, proteínas transportadoras entre células do mesofilo e bainha do feixe vascular, além de inúmeras enzimas catalíticas que sustentam o metabolismo vegetal como um todo (Taiz et al., 2017; Foyer & Noctor, 2011).
Dessa forma, compreender a integração entre metabolismo fotossintético C4 e metabolismo do nitrogênio é fundamental para o manejo agronômico da cultura. A eficiência no uso do nitrogênio não depende apenas da quantidade ofertada no solo, mas da capacidade fisiológica da planta em converter esse nutriente em estrutura fotossintética, metabolismo ativo e produção de biomassa e grãos. Nesse contexto, o nitrogênio deixa de ser apenas um nutriente e passa a ser componente central da construção da alta performance fisiológica do milho.
A aquisição desse macronutriente ocorre por diferentes vias. O nitrogênio pode ser disponibilizado pela mineralização da matéria orgânica do solo, pela aplicação de fertilizantes nitrogenados e também pela contribuição de microrganismos promotores de crescimento, como bactérias do grupo dos Azospirillum (Robertson & Vitousek, 2009; Kuypers et al., 2018; Hungria et al., 2010; Fukami et al., 2018).
Isso reforça um ponto importante no manejo agronômico. Estratégias que favoreçam a absorção, a assimilação e a eficiência metabólica do nitrogênio são determinantes para que a cultura expresse seu máximo potencial produtivo.
Como ocorre a entrada de nitrogênio no sistema produtivo
O nitrogênio presente no sistema agrícola não está disponível em uma única forma. Sua dinâmica varia conforme as condições físico-químicas do solo e a atividade dos microrganismos. Nesse sentido, é importante entender quais são as principais formas de nitrogênio presentes na solução do solo e como isso influencia diretamente a absorção pela planta para se obter mais eficiência no seu uso.
O estoque de nitrogênio nativo do solo é formado, principalmente, pela incorporação do N₂ atmosférico via fixação biológica, sendo está a principal via de entrada de nitrogênio novo no sistema. Esse processo é realizado por bactérias diazotróficas que atuam em diferentes formas, incluindo associações simbióticas, associativas e de vida livre no solo.
Embora sistemas simbióticos apresentem maior eficiência, especialmente em leguminosas, a fixação biológica não se restringe a esses sistemas, ocorrendo também em outras interações planta-microrganismo e diretamente no ambiente edáfico (Liu et al., 2022; Liu et al., 2024).
Do ponto de vista bioquímico, o N₂ atmosférico é inicialmente reduzido a amônia (NH₃) pela ação da nitrogenase, um complexo enzimático dependente de cofatores metálicos, principalmente ferro (Fe) e molibdênio (Mo), organizados no FeMo-cofactor. Essa reação envolve transferência sequencial de elétrons e elevado consumo energético, com gasto aproximado de 16 moléculas de ATP por molécula de N₂ fixada. No ambiente celular, a NH₃ rapidamente se protona, formando amônio (NH₄⁺), que constitui a primeira forma inorgânica assimilável do nitrogênio proveniente da fixação biológica (Rutledge e Tezcan, 2023).
Independentemente da forma de associação, o nitrogênio fixado, na forma de NH₄⁺, pode ser assimilado pelos microrganismos para seu próprio metabolismo. Em sistemas com interação planta-microrganismo, o NH₄⁺ é transferido à planta, onde é incorporado ao metabolismo por meio da síntese de aminoácidos (Zayed et al., 2023; Xu et al., 2012).
Posteriormente, esse nitrogênio retorna ao solo por meio da decomposição de resíduos culturais, exsudação radicular e turnover de raízes e estruturas microbianas, passando a compor o pool de nitrogênio orgânico do solo. A partir desse reservatório, processos de mineralização convertem o nitrogênio orgânico novamente em formas inorgânicas, como NH₄⁺ e NO₃⁻, fechando o ciclo e determinando a disponibilidade de nitrogênio na solução do solo (Udvardi e Poole, 2013; Liu et al., 2024).
Plantas de cobertura e eficiência do nitrogênio no milho
A dinâmica de mineralização e disponibilização do nitrogênio no solo está diretamente relacionada à qualidade e à quantidade de resíduos orgânicos incorporados ao sistema, sendo as plantas de cobertura um dos principais fatores de modulação desse processo.
O tipo de planta de cobertura exerce influência significativa sobre a dinâmica do nitrogênio no solo. Em estudos realizados por Silva et al. (2006), verificou-se que a crotalária, por ser uma leguminosa, contribui com maior aporte de nitrogênio ao sistema, enquanto o milheto apresenta papel mais relacionado à ciclagem e conservação, com menor fornecimento líquido de nitrogênio. Além disso, os autores observaram maior produtividade do milho no sistema crotalária-milho, em função do maior aporte de nitrogênio durante a mineralização dos resíduos culturais da crotalária.
A crotalária (Crotalaria juncea), por ser uma leguminosa altamente eficiente na fixação biológica de nitrogênio, apresenta elevado potencial de aporte ao sistema. Nery e Franco Junior (2023) verificaram um aporte de 40,95 kg ha⁻¹ de nitrogênio ao utilizar a C. juncea como planta de cobertura. Além disso, os autores destacam que essa espécie promove elevada produção de biomassa, contribuindo para a melhoria das propriedades físico-químicas e biológicas do solo, sendo considerada um adubo verde estratégico para sistemas sustentáveis.
As plantas de cobertura Urochloa ruziziensis (braquiária), Guizotia abyssinica (niger) e Crambe abyssinica (crambe) incorporam valores de 6,48 kg ha⁻¹, 29,72 kg ha⁻¹ e 15,10 kg ha⁻¹ de nitrogênio, respectivamente (Nery e Franco Junior, 2023).
Resultados semelhantes foram observados por Ambrosano et al. (2006), que verificaram que a crotalária, na média de dois anos agrícolas, proporcionou ao milho cultivado sem adubação nitrogenada um efeito equivalente à aplicação de 56,0 e 73,0 kg ha⁻¹ de N-ureia, em comparação ao milho cultivado em pousio e em sucessão ao milheto.
É importante ressaltar que, apesar da crotalária apresentar maior eficiência no fornecimento direto de nitrogênio ao sistema, as gramíneas, como milheto e braquiária, desempenham papel fundamental na sustentabilidade do sistema produtivo. Essas espécies promovem a ciclagem de nutrientes, proteção do solo, incremento da matéria orgânica e melhoria das propriedades físicas, químicas e biológicas do solo.
Como o manejo das plantas de cobertura influencia o nitrogênio do solo e sua disponibilidade para o milho
As plantas de cobertura influenciam diretamente a dinâmica das formas de nitrogênio no solo durante seu crescimento, principalmente pela absorção e retenção de nitrogênio mineral, com destaque para o NO₃⁻, reduzindo perdas por lixiviação. Além disso, espécies com sistema radicular profundo, como Urochloa spp., exploram camadas mais profundas do solo, promovendo a recuperação de nitrogênio lixiviado e sua redistribuição no sistema, aumentando a eficiência da ciclagem de nutrientes (Boddey et al., 2010). Adicionalmente, também modulam a atividade microbiana e a ciclagem de nutrientes, influenciando a disponibilidade de N no sistema (Abdalla et al., 2019).
Esse efeito é mais evidente em gramíneas, que atuam como espécies conservadoras de nitrogênio, mantendo-o temporariamente imobilizado na biomassa vegetal (Justes et al., 2012). Dessa forma, durante o desenvolvimento das plantas de cobertura, ocorre maior retenção de N no sistema, reduzindo perdas e favorecendo a eficiência do uso de nitrogênio pelo milho em sucessão (Carvalho et al., 2024).
Após a dessecação e decomposição dos resíduos, a dinâmica do nitrogênio passa a ser regulada pela mineralização da matéria orgânica, processo fortemente influenciado pela qualidade dos resíduos, especialmente pela relação C:N.
Espécies com baixa relação C:N, como leguminosas, favorecem rápida liberação de NH₄⁺, posteriormente convertido em NO₃⁻, aumentando sua disponibilidade ao milho. Por outro lado, gramíneas promovem liberação mais gradual, em função da maior imobilização microbiana (Abdalla et al., 2019) (Figura 3). Assim, a escolha da planta de cobertura e seu manejo determinam a sincronização entre a liberação de nitrogênio e a demanda da cultura do milho, sendo esse um dos principais fatores para maximizar a eficiência do uso de N no sistema produtivo (Carvalho et al., 2024).

Como o nitrogênio se transforma no solo e fica disponível para as plantas
O nitrogênio no solo está em constante transformação entre formas orgânicas e inorgânicas, sendo a matéria orgânica seu principal reservatório. A partir desse reservatório , microrganismos promovem a mineralização, convertendo o N orgânico em NH₄⁺, processo diretamente dependente da atividade biológica e das condições do ambiente (Ros et al., 2011; Grzyb et al., 2021).
Em condições aeradas, o NH₄⁺ é rapidamente transformado em NO₃⁻ pela nitrificação, mediada por microrganismos específicos (figura 4). Dessa forma, a predominância entre NH₄⁺ e NO₃⁻ no solo passa a ser definida principalmente pela atividade microbiana e pelas condições físico-químicas do ambiente (Huang et al., 2021).
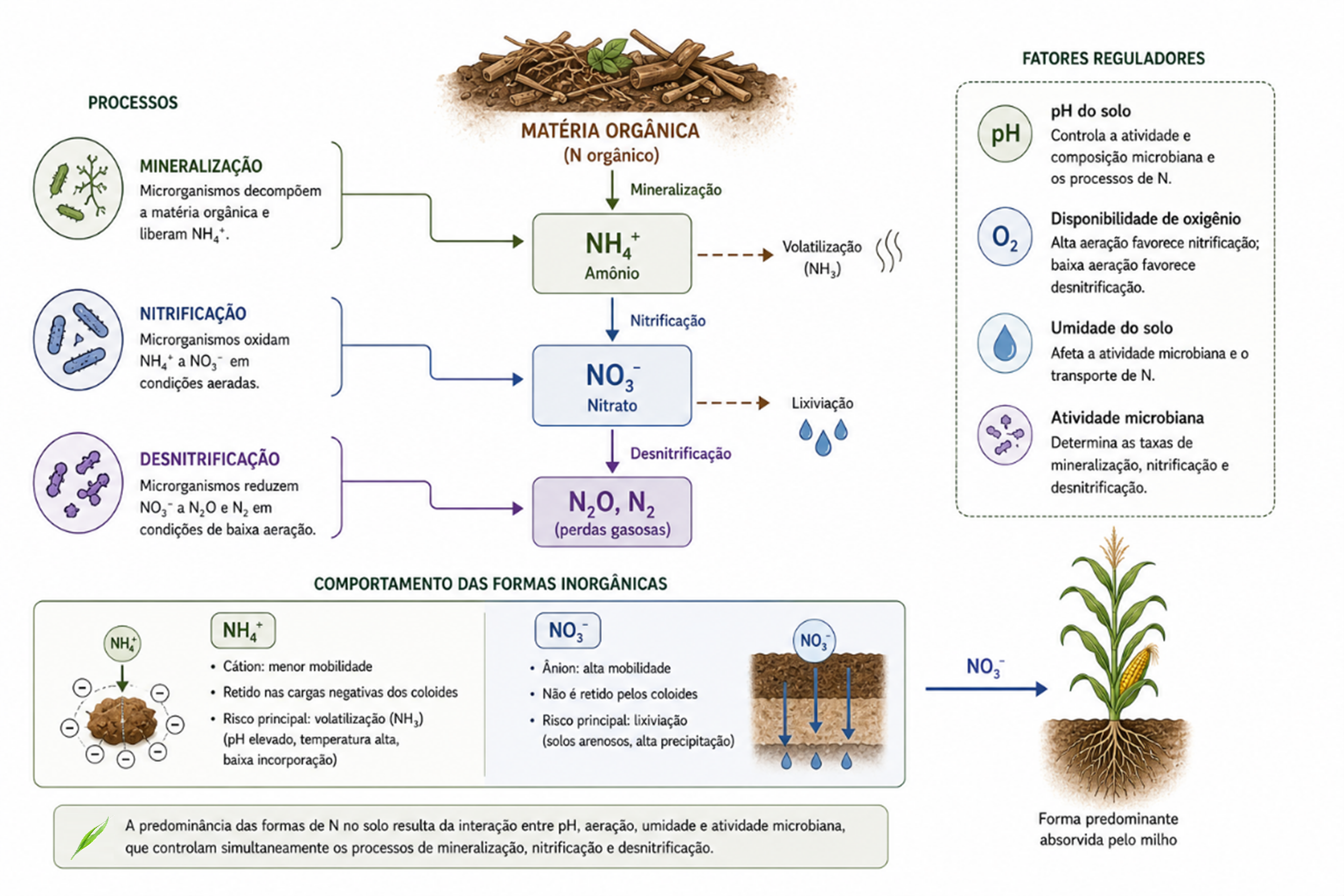
O pH do solo exerce papel central nesse processo. Em pH próximo à neutralidade, há maior atividade nitrificante e maior formação de NO₃⁻. Já em solos ácidos, a nitrificação pode ser reduzida. Em pH elevado, aumenta o risco de perdas por volatilização de NH₃ (Qiu et al., 2024; Grzyb et al., 2021).
A aeração do solo também é determinante. Em solos bem aerados, predominam processos oxidativos, favorecendo a formação de NO₃⁻. Em condições de baixa aeração, como em solos compactados ou encharcados, ocorre desnitrificação, com redução do NO₃⁻ a N₂O e N₂, resultando em perdas de nitrogênio (Qiu et al., 2024).
Do ponto de vista prático, o NH₄⁺ apresenta menor mobilidade, podendo ser retido no solo, mas com risco de volatilização. Já o NO₃⁻ é altamente móvel, sujeito à lixiviação, principalmente em solos arenosos e sob alta precipitação (Grzyb et al., 2021; Carvalho et al., 2024).
Assim, a disponibilidade de nitrogênio no sistema resulta da interação entre pH, aeração, umidade e atividade microbiana. No milho, que absorve predominantemente NO₃⁻, o manejo deve buscar sincronizar a formação dessa forma com a demanda da cultura, reduzindo perdas e aumentando a eficiência do uso do nitrogênio (Carvalho et al., 2024).
Como o milho regula a absorção de nitrogênio: papel dos transportadores e das formas de N
A absorção de nitrogênio pelo milho ocorre predominantemente na forma de nitrato (NO₃⁻), refletindo, em primeiro lugar, a dinâmica do nitrogênio no solo. Em sistemas tropicais, com boa aeração e alta atividade microbiana, o NH₄⁺ oriundo da mineralização ou da adubação é rapidamente convertido em NO₃⁻ pela nitrificação, tornando o nitrato a forma mais disponível no ambiente radicular (Grzyb et al., 2021; Huang et al., 2021).
No entanto, a absorção não depende apenas da disponibilidade no solo (Figura 5). A planta exerce forte controle por meio de transportadores específicos. O NO₃⁻ é absorvido por sistemas de alta (NRT2) e baixa afinidade (NRT1), que se ajustam conforme a concentração no solo. Em baixa disponibilidade, predominam transportadores de alta afinidade, aumentando a eficiência de captação. Em maior disponibilidade, ocorre maior fluxo via transportadores de baixa afinidade (Dechorgnat et al., 2019).

Além disso, o nitrato atua como molécula sinalizadora. O transportador NRT1.1 (NPF6.3) integra transporte e sinalização, regulando a expressão de genes ligados à absorção e assimilação de nitrogênio, ajustando a resposta da planta às condições do ambiente (Undurraga et al., 2017).
Essa regulação também depende do estado nutricional interno. A glutamina, como indicador do status nitrogenado, exerce feedback negativo sobre a absorção em condições de suficiência e estimula a expressão de transportadores em situações de deficiência (Dechorgnat et al., 2019; Masclaux-Daubresse et al., 2010).
Para o NH₄⁺, a absorção ocorre via transportadores da família AMT, sob controle mais restritivo. Como o amônio não pode se acumular no citoplasma, ele precisa ser rapidamente assimilado via GS/GOGAT. Quando a absorção excede essa capacidade, ocorre acúmulo e risco de toxicidade, levando à redução da atividade dos transportadores como mecanismo de proteção (Britto e Kronzucker, 2002; 2013; Lea e Miflin, 2012).
Do ponto de vista do manejo, a aplicação de fontes amoniacais aumenta temporariamente a disponibilidade de NH₄⁺ e sua absorção inicial. No entanto, em solos bem aerados, a rápida nitrificação converte esse amônio em NO₃⁻, restabelecendo o nitrato como principal forma absorvida pela cultura.
Assim, a absorção de nitrogênio pelo milho resulta da interação entre a forma predominante no solo e o controle fisiológico da planta. Na prática, isso reforça que o manejo deve considerar não apenas a dose, mas a forma de N aplicada e as condições do solo, buscando sincronizar a disponibilidade com a capacidade de absorção da cultura para maximizar a eficiência do uso do nitrogênio.
Fontes de nitrogênio na adubação do milho
A dinâmica das formas de nitrogênio no solo, caracterizada pela rápida conversão entre NH₄⁺ e NO₃⁻, estabelece a base para compreender o comportamento das diferentes fontes nitrogenadas na adubação do milho.
Embora a cultura absorva nitrogênio predominantemente na forma de nitrato, as fontes diferem quanto à forma inicial de disponibilização e, principalmente, quanto à suscetibilidade às perdas no sistema solo-atmosfera.
No Brasil, a ureia é a principal fonte utilizada, representando cerca de 55–65% do consumo de fertilizantes nitrogenados, seguida pelo sulfato de amônio, com participação entre 15–25% (ANDA, 2023; IFA, 2022). Essa predominância está associada à sua elevada concentração de nitrogênio (45–46%), o que reduz custos de transporte e aplicação por unidade de nutriente, além de apresentar menor custo por kg de N em comparação às demais fontes, favorecendo sua adoção em larga escala.
Do ponto de vista experimental, Cantarella et al. (2007) demonstram que a ureia, quando aplicada superficialmente, apresenta elevadas perdas por volatilização de NH₃, reduzindo a recuperação do nitrogênio pela cultura. Esses resultados são consistentes com observações posteriores em sistemas tropicais, nas quais a volatilização se configura como uma das principais vias de perda de nitrogênio quando não há incorporação ou precipitação após a aplicação (EMBRAPA, 2019).
Resultados comparativos reforçam essa dinâmica. Lara Cabezas et al. (2000) observaram maior produtividade do milho com o uso de sulfato de amônio em relação à ureia em condições de aplicação superficial.
De forma semelhante, Ernani et al. (2013) demonstram que fontes menos suscetíveis à volatilização apresentam maior eficiência de uso do nitrogênio em sistemas de plantio direto.
Por outro lado, Meira et al. (2009), ao comparar ureia, sulfato de amônio e nitrato de amônio, observaram ausência de diferença significativa na produtividade do milho quando houve condições favoráveis à incorporação ou ocorrência de chuva após a aplicação.
Assim, os resultados experimentais convergem para um ponto central: as diferenças entre fontes nitrogenadas estão fortemente associadas às perdas no sistema solo-atmosfera.
A ampla utilização da ureia no Brasil reflete sua vantagem econômica e operacional, enquanto sua eficiência agronômica depende diretamente do manejo.
Dessa forma, a escolha da fonte deve considerar o ambiente de aplicação e o risco de perdas, priorizando estratégias que garantam a permanência do nitrogênio no sistema até o momento de sua absorção pela cultura.
Manejo da adubação nitrogenada no milho
A eficiência do manejo da adubação nitrogenada no milho depende da correta compreensão da demanda da cultura e da definição adequada das doses aplicadas. De forma geral, o milho necessita entre 20 e 25 kg de N para cada tonelada de grãos produzida (Bender et al., 2013).
Em condições brasileiras, trabalhos experimentais indicam respostas consistentes da cultura ao aumento das doses de nitrogênio até faixas entre 120 e 180 kg de N ha⁻¹, dependendo do sistema de produção, nível de produtividade e disponibilidade de nitrogênio no solo (Figura 6) (EMBRAPA, 2013; Cantarella et al., 2007).

A relação entre dose e produtividade segue uma curva típica de resposta, com aumento de rendimento até um ponto de máxima eficiência agronômica, seguido por estabilização.
Estudos mostram que, em muitos ambientes tropicais, a máxima eficiência agronômica ocorre entre 120 e 150 kg de N ha⁻¹, faixa em que há maior retorno produtivo por unidade de N aplicado. Acima desse intervalo, embora ainda possa haver incremento de produtividade, ocorre redução da eficiência do uso do nitrogênio, caracterizando o início da zona de saturação do sistema (EMBRAPA, 2013; Bender et al., 2013).
Doses superiores a 180–200 kg de N ha⁻¹, em grande parte dos experimentos conduzidos no Brasil, apresentam baixo retorno adicional em produtividade e aumento expressivo do risco de perdas por volatilização, lixiviação e desnitrificação. Esse comportamento indica que a limitação do sistema deixa de ser a disponibilidade de nitrogênio e passa a ser fisiológica ou ambiental, reduzindo a eficiência agronômica do fertilizante aplicado (Cantarella et al., 2007; EMBRAPA, 2013).
Essa definição de dose deve ser integrada à marcha de absorção da cultura. Até V4–V6, o milho absorve aproximadamente 20–30% do nitrogênio total requerido, com a maior parte da absorção ocorrendo entre V6 e o florescimento (Plenet e Lemaire, 2000; Bender et al., 2013).
Nesse contexto, trabalhos experimentais demonstram que a aplicação de nitrogênio em cobertura entre V3 e V6 maximiza a eficiência do uso do nutriente, por alinhar a disponibilidade de N com o início da fase de maior demanda da cultura (EMBRAPA, 2013).
Assim, o manejo eficiente da adubação nitrogenada no milho depende da integração entre dose e momento de aplicação. Doses abaixo da faixa ótima limitam o potencial produtivo, enquanto doses acima do ponto de máxima eficiência aumentam perdas e reduzem o retorno agronômico.
Dessa forma, a máxima eficiência do sistema é alcançada quando a dose é ajustada ao potencial produtivo e o fornecimento de nitrogênio é sincronizado com a fase de maior absorção da cultura, garantindo maior aproveitamento do nutriente e estabilidade produtiva.
Como aumentar a eficiência do nitrogênio no milho
Após a adubação nitrogenada, ocorre aumento expressivo da concentração de nitrato (NO₃⁻) na solução do solo, resultado da rápida nitrificação do amônio em sistemas tropicais. Em estádios iniciais da cultura, especialmente até V4–V6, a planta ainda apresenta baixa demanda relativa por nitrogênio, favorecendo a permanência do nutriente no sistema e aumentando o risco de perdas por lixiviação (Huang et al., 2021).
Nesse contexto, a eficiência do uso do nitrogênio depende diretamente da capacidade da planta em absorver e, principalmente, assimilar rapidamente o nitrato disponibilizado.
A assimilação do nitrato inicia-se com a redução do nitrato (NO₃⁻) a nitrito (NO₂⁻) pela enzima redutase do nitrato (NR), localizada no citosol e dependente de NADH ou NADPH como fonte de elétrons, tendo o molibdênio como cofator essencial (Kaiser et al., 2011).
A limitação de molibdênio reduz a atividade da redutase do nitrato, promovendo o acúmulo de nitrato nos tecidos e redução na taxa de assimilação do nitrogênio.
Em milho, a aplicação de molibdênio está associada ao aumento da assimilação de nitrogênio e ao incremento no rendimento de grãos, com produtividades elevando-se de aproximadamente 5,8 para 6,6 t ha⁻¹, evidenciando seu papel na eficiência do uso do nitrogênio (Imran et al., 2020).
Na sequência, o nitrito é reduzido a amônio (NH₄⁺) pela enzima redutase do nitrito (NiR), nos plastídios, utilizando ferredoxina reduzida como fonte de elétrons, em um processo diretamente dependente da atividade fotossintética (Xu et al., 2020).
O amônio gerado é então assimilado via sistema glutamina sintetase / glutamato sintase (GS/GOGAT), no qual a glutamina sintetase incorpora NH₄⁺ ao glutamato com consumo de ATP, formando glutamina, e a glutamato sintase utiliza poder redutor e o esqueleto de carbono 2-oxoglutarato para formar glutamato (Nunes-Nesi et al., 2021). Esse processo representa o principal custo energético da assimilação do nitrogênio (Figura 6).
No milho, essa integração é favorecida pelo metabolismo fotossintético C4, que aumenta a disponibilidade de ATP, poder redutor (ferredoxina/NADPH) e esqueletos de carbono derivados do metabolismo central, especialmente o 2-oxoglutarato. Essa condição favorece a assimilação do nitrogênio absorvido e reduz o acúmulo de formas minerais nos tecidos, principalmente após aplicações de fontes nitrogenadas amoniacais, nas quais ocorre aumento transitório da absorção de NH₄⁺ (Nunes-Nesi et al., 2021).

Além da assimilação, a eficiência do uso do nitrogênio depende da reciclagem interna do nutriente. Nesse processo, o níquel atua como cofator da enzima urease, responsável pela hidrólise da ureia e liberação de amônio (NH₄⁺) no metabolismo da planta.
A disponibilidade de níquel (Ni) regula diretamente a atividade da urease e o fluxo de reciclagem do nitrogênio, sendo sua deficiência associada ao acúmulo de ureia e menor eficiência metabólica do nutriente (Witte et al., 2011).
Em milho, a aplicação de níquel na dose de 25 mg kg⁻¹ de solo aumentou a atividade da urease, promoveu maior absorção e acúmulo de nitrogênio na planta e elevou a biomassa, com incremento de 24% na massa seca da parte aérea, além de aumento no teor de nitrogênio foliar em relação ao controle (Uğurlar, 2024).
Em condições de campo, Gomes et al. (2025) demonstraram que a aplicação foliar de níquel na dose de 80 g ha⁻¹ aumentou a taxa fotossintética, o teor de clorofila e resultou em incremento significativo no rendimento de grãos em relação ao tratamento sem aplicação, evidenciando seu papel na eficiência do uso do nitrogênio na cultura do milho.
Em condições de elevada disponibilidade de nitrogênio, especialmente após aplicações em estádios iniciais, pode ocorrer acúmulo transitório de compostos nitrogenados, resultando em desequilíbrio metabólico e aumento da síntese de etileno. O cobalto atua como modulador desse processo ao inibir a conversão de ACC em etileno, contribuindo para maior estabilidade fisiológica da planta (Taiz et al., 2017; Farooq et al., 2022).
Em milho, a aplicação de cobalto em baixas doses promoveu aumento do crescimento vegetativo, incluindo maior altura de plantas, expansão da área foliar e incremento no número de espigas por planta, refletindo em aumento nos componentes de produção (Jayakumar et al., 2015).
Em condições de estresse hídrico, sua aplicação também foi associada ao aumento da produtividade e da eficiência no uso da água (Khalifa et al., 2023). Por outro lado, concentrações elevadas de cobalto podem reduzir o crescimento e a atividade fotossintética, evidenciando resposta dependente da dose (Anjum et al., 2023).
Assim, a eficiência do uso de nitrogênio no milho resulta da integração entre disponibilidade no sistema, capacidade de assimilação e regulação metabólica. O molibdênio atua diretamente na assimilação do nitrato, o níquel na reciclagem interna do nutriente e o cobalto como modulador fisiológico, permitindo maior conversão do nitrogênio em formas orgânicas, redução de perdas e aumento da eficiência produtiva da cultura.
Conclusões finais
A alta eficiência do milho não é explicada apenas pelo metabolismo fotossintético C4, mas pela forte integração entre os metabolismos do carbono e do nitrogênio. O sistema C4 aumenta a eficiência fotossintética, reduz a fotorespiração e fornece energia e esqueletos de carbono essenciais para a assimilação do nitrogênio.
Além disso, o nitrogênio participa diretamente da formação da maquinaria fotossintética da planta, compondo clorofilas, enzimas e proteínas do transporte de elétrons. Compostos como glutamato e aspartato conectam o metabolismo do carbono à síntese de aminoácidos e proteínas, sustentando o crescimento vegetal.
Dessa forma, a elevada produtividade do milho resulta da coordenação entre fotossíntese, absorção e assimilação de nitrogênio. Por isso, a eficiência no uso do N depende não apenas da dose aplicada, mas da capacidade fisiológica da planta em transformar esse nutriente em metabolismo ativo, biomassa e produção de grãos.
Aprofunde seu manejo com conteúdo técnico
Para apoiar decisões de campo e estruturar um manejo integrado mais eficiente, a Agroadvance baixe o e-Book completo sobre Nutrição e Proteção de Soja e Milho. É gratuito. Basta clicar em SAIBA MAIS!


Dê o próximo passo na sua carreira
Conheça a Pós-Graduação em Soja e Milho da Agroadvance. O curso conecta fundamentos científicos, tecnologia agrícola e interpretação agronômica para formar profissionais capazes de tomar decisões precisas em ambientes complexos, que vão da proteção de plantas ao manejo nutricional, da fisiologia ao entendimento epidemiológico de doenças como a mancha-alvo. Clique em SAIBA MAIS e conheça!

Referências
ABDALLA, M. et al. Conservation agriculture effects on soil organic carbon and nitrogen dynamics. Agriculture, Ecosystems & Environment, v. 279, p. 33–45, 2019.
AMBROSANO, E. J. et al. Nitrogen supply to maize from leguminous cover crops. Scientia Agricola, v. 63, n. 6, p. 547–553, 2006.
ANJUM, S. A. et al. Cobalt toxicity in plants: physiological and biochemical responses. Environmental and Experimental Botany, v. 211, p. 105319, 2023.
ARRIVAULT, S. et al. Metabolite profiles reveal the intercellular metabolite exchange underlying C4 photosynthesis in maize. Plant Cell, v. 29, n. 5, p. 1076–1091, 2017. DOI: 10.1093/jxb/erw414
AVILA, R. G. et al. Drought-tolerant maize genotypes invest in root system and maintain high harvest index during water stress. Revista Brasileira de Milho e Sorgo, v. 15, n. 3, p. 450–460, 2016.
BENDER, R. R. et al. Nutrient uptake, partitioning, and remobilization in modern maize hybrids. Agronomy Journal, v. 105, p. 161–170, 2013.
BLOOM, A. J. The increasing importance of distinguishing among plant nitrogen sources. Current Opinion in Plant Biology, v. 25, p. 10–16, 2015.
BODDEY, R. M. et al. Biological nitrogen fixation in tropical grasses. Plant and Soil, v. 334, p. 1–18, 2010.
BRITTO, D. T.; KRONZUCKER, H. J. NH₄⁺ toxicity in plants: a review. Journal of Plant Physiology, v. 159, p. 567–584, 2002.
CAMPOS, C. N.; ÁVILA, R. G.; SOUZA, K. R. D.; AZEVEDO, L. M.; ALVES, J. D. Melatonin reduces oxidative stress and promotes drought tolerance in young Coffea arabica L. plants. Agricultural Water Management, v. 211, p. 37–47, 2019.
CANTARELLA, H. et al. Ammonia volatilization from nitrogen fertilizers. Revista Brasileira de Ciência do Solo, v. 31, p. 491–501, 2007.
CARVALHO, J. L. N. et al. Nitrogen cycling and efficiency in tropical cropping systems. Agronomy, v. 14, p. 215, 2024.
DECHORGNAT, J. et al. Nitrate transporters in plants. Journal of Experimental Botany, v. 70, p. 2451–2465, 2019.
EDWARDS, G. E.; FRANCESCHI, V. R.; VOZNESENSKAYA, E. V. Single-cell C4 photosynthesis versus the dual-cell paradigm. Annual Review of Plant Biology, v. 55, p. 173–196, 2004.
EMBRAPA. Adubação nitrogenada na cultura do milho. Brasília: Embrapa, 2013.
EMBRAPA. Boas práticas para uso eficiente de fertilizantes nitrogenados. Brasília: Embrapa, 2019.
ERNANI, P. R. et al. Nitrogen sources and efficiency in no-tillage systems. Revista Brasileira de Ciência do Solo, v. 37, p. 171–178, 2013.
ERMAKOVA, M. et al. On the road to C4 rice: advances and perspectives. Plant Journal, v. 101, p. 940–950, 2020.
EVANS, J. R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia, v. 78, p. 9–19, 1989.
FAROOQ, M. et al. Role of cobalt in plant stress physiology. Plants, v. 11, p. 1234, 2022.
FOYER, C. H.; NOCTOR, G. Ascorbate and glutathione: the heart of the redox hub. Plant Physiology, v. 155, p. 2–18, 2011.
FOYER, C. H. et al. Coordination of carbon and nitrogen metabolism in plants. Journal of Experimental Botany, v. 62, p. 1399–1412, 2011.
FUKAMI, J.; CEREZINI, P.; HUNGRIA, M. Azospirillum: benefits that go far beyond biological nitrogen fixation. AMB Express, v. 8, p. 73, 2018.
FURBANK, R. T. Evolution of the C4 photosynthetic mechanism. Journal of Experimental Botany, v. 62, p. 3103–3108, 2011.
FURBANK, R. T. Walking the C4 pathway: past, present, and future. Journal of Experimental Botany, v. 67, p. 4057–4066, 2016.
GHANNOUM, O. et al. Improving photosynthesis and crop productivity. Plant Physiology, v. 155, p. 194–205, 2011.
GOMES, A. L. et al. Nickel foliar application and maize yield response. Agronomy, v. 15, p. 112, 2025.
GRZYB, A. et al. Nitrogen transformations in agricultural soils. Soil Biology and Biochemistry, v. 153, p. 108112, 2021.
HATCH, M. D. C4 photosynthesis. Biochimica et Biophysica Acta, v. 895, p. 81–106, 1987.
HATCH, M. D.; BURNELL, J. N. Carbonic anhydrase activity in leaves. Plant Physiology, v. 93, p. 825–828, 1990.
HUANG, M. et al. Nitrification and nitrogen cycling in soils. Science of the Total Environment, v. 753, p. 141708, 2021.
HUNGRIA, M. et al. Inoculation with Azospirillum brasilense improves maize yield. Plant and Soil, v. 331, p. 413–425, 2010.
IFA. Fertilizer consumption report. Paris: International Fertilizer Association, 2022.
IMRAN, M. et al. Role of molybdenum in nitrogen metabolism. Agronomy, v. 10, p. 156, 2020.
JAYAKUMAR, K. et al. Effect of cobalt on maize growth and yield. International Journal of Agricultural Science, v. 7, p. 45–52, 2015.
JUSTES, E. et al. Cover crops and nitrogen cycling. Agronomy for Sustainable Development, v. 32, p. 35–50, 2012.
KAISER, W. M. et al. Nitrate reductase regulation. Journal of Experimental Botany, v. 62, p. 2299–2311, 2011.
KHALIFA, G. S. et al. Cobalt improves drought tolerance in maize. Plant Physiology Reports, v. 28, p. 220–231, 2023.
KUYPERS, M. M. M. et al. The microbial nitrogen-cycling network. Nature Reviews Microbiology, v. 16, p. 263–276, 2018.
LARA CABEZAS, W. A. R. et al. Nitrogen sources and application methods in maize. Scientia Agricola, v. 57, p. 177–182, 2000.
LEA, P. J.; MIFLIN, B. J. Nitrogen assimilation. Annals of Botany, v. 109, p. 135–152, 2012.
LIU, X. et al. Biological nitrogen fixation. Global Change Biology, v. 28, p. 1453–1467, 2022.
LIU, X. et al. Nitrogen cycling in soil-plant systems. Nature Reviews Earth & Environment, v. 5, p. 120–134, 2024.
MASCLAUX-DAUBRESSE, C. et al. Nitrogen metabolism and recycling. Annual Review of Plant Biology, v. 61, p. 369–393, 2010.
MEIRA, F. A. et al. Nitrogen sources and maize productivity. Revista Brasileira de Milho e Sorgo, v. 8, p. 115–124, 2009.
MU, X. et al. Photosynthetic nitrogen-use efficiency in maize. Plant Physiology, v. 172, p. 396–408, 2016.
NERY, A. R.; FRANCO JUNIOR, K. S. Cover crops and nitrogen contribution. Agronomy Journal, v. 115, p. 112–121, 2023.
NUNES-NESI, A. et al. Carbon–nitrogen interactions in plants. Molecular Plant, v. 3, p. 973–996, 2010.
NUNES-NESI, A. et al. Integration of carbon and nitrogen metabolism. Plant Physiology, v. 187, p. 159–173, 2021.
PLENET, D.; LEMAIRE, G. Nitrogen uptake in maize. Agronomy Journal, v. 92, p. 521–527, 2000.
QIU, S. et al. Soil pH and nitrogen cycling. Soil Biology and Biochemistry, v. 185, p. 108812, 2024.
ROBERTSON, G. P.; VITOUSEK, P. M. Nitrogen in agriculture. Annual Review of Environment and Resources, v. 34, p. 97–125, 2009.
ROS, G. H. et al. Soil nitrogen mineralization. Soil Biology and Biochemistry, v. 43, p. 1049–1058, 2011.
RUTLEDGE, H. L.; TEZCAN, F. A. Nitrogenase mechanism. Chemical Reviews, v. 123, p. 12345–12378, 2023.
SAGE, R. F. The evolution of C4 photosynthesis. New Phytologist, v. 161, p. 341–370, 2004.
SAGE, R. F.; KUBIEN, D. S. Temperature response of C3 and C4 photosynthesis. Plant, Cell & Environment, v. 30, p. 1086–1106, 2007.
SAGE, R. F.; ZHU, X. G. Exploiting the engine of C4 photosynthesis. Journal of Experimental Botany, v. 62, p. 2989–3000, 2011.
SINCLAIR, T. R.; HORIE, T. Leaf nitrogen and photosynthesis. Crop Science, v. 29, p. 90–98, 1989.
STUDER, A. J. et al. Genetic control of carbonic anhydrase in maize. Plant Physiology, v. 164, p. 1505–1515, 2014. DOI: 10.1104/pp.114.237602
TAIZ, L. et al. Fisiologia e desenvolvimento vegetal. Porto Alegre: Artmed, 2017.
UNDURRAGA, S. F. et al. Dual function of NRT1.1 transporter. Plant Cell, v. 29, p. 243–256, 2017.
UDVARDI, M.; POOLE, P. S. Plant–microbe interactions. Annual Review of Plant Biology, v. 64, p. 781–805, 2013.
VON CAEMMERER, S.; FURBANK, R. T. Improving C4 photosynthesis. Current Opinion in Plant Biology, v. 31, p. 125–134, 2016.
WINGLER, A. et al. Photorespiration and stress. Philosophical Transactions of the Royal Society B, v. 355, p. 1517–1529, 1999.
WITTE, C. P. et al. Urease and nitrogen recycling. Plant Biology, v. 13, p. 1–9, 2011.
XU, G. et al. Nitrogen assimilation. Annual Review of Plant Biology, v. 63, p. 153–182, 2012.
XU, G. et al. Nitrogen metabolism signaling. Plant Communications, v. 1, p. 100014, 2020.
YAMORI, W. et al. Temperature response of photosynthesis. Photosynthesis Research, v. 119, p. 101–117, 2014.
ZAYED, A. et al. Nitrogen assimilation and plant–microbe interaction. Frontiers in Plant Science, v. 14, p. 115432, 2023.
UĞURLAR, F. Nickel effects on nitrogen metabolism in maize. Journal of Plant Nutrition, v. 47, p. 1–12, 2024.
Sobre o autor:

Como citar este artigo:
AVILA, R.G. Nitrogênio no milho: como aumentar a eficiência do nutriente e a produtividade da lavoura. Blog Agroadvance. Publicado em: 18 Mai. 2026. Disponível em: https://agroadvance.com.br/blog-nitrogenio-no-milho/. Data de acesso: 21 jul. 2026




